After the 2010 Nowak, Tarnita, & Wilson affair, Samir Okasha weighed in with a comment in Nature, effectively asking: why can't we all just get along?
The paper was titled: Altruism researchers must cooperate. It is indeed ironic that the science of cooperation has led to so much scientific acrimony.
In the paper, Samir embraces equivalence, writing:
kin and multi-level selection are not alternative theories; they simply offer different takes on the question of how social behaviour evolved. Proponents of kin selection, for example, explain sterile workers in insect colonies by saying that the workers are helping the queen to reproduce, and thus boosting their own inclusive fitness. Proponents of multi-level selection argue that the workers are providing a benefit to the colony as a whole, thus making the colony fitter than other colonies. These explanations may seem different, but mathematical models show that they are in fact equivalent
He says the persistent rival camps are a puzzle since:
The existence of equivalent formulations of a theory, or of alternative modelling approaches, does not usually lead to rival camps in science. The Lagrangian and Hamiltonian formulations of classical mechanics, for example, or the wave and matrix formulations of quantum mechanics, tend to be useful for tackling different problems, and physicists switch freely between them.
His explanation:
History shows that, despite its enormous empirical success, evolutionary biology is peculiarly susceptible to controversy and infighting. This is particularly true of social evolution theory, in part because of its potential applications to human behaviour.
I think this is essentially correct. People fight over application of evolutionary theory to humans for many reasons, but among them are: "it's complicated", and "it's important to get it right".
Group selection doesn't score well in the latter category. It isn't so much that it is wrong, it's more that it has led to decades of muddle, confusion and poor-quality science. If kin selection and group selection are feeling different parts of the same elephant, kin selection has hold of the trunk, while group selection is groping the left thigh.
It is certainly frustrating to have all the biology papers using kin selection, while all the humanities papers seem to use group selection.
A big part of the problem is misunderstandings surrounding cultural evolution. Cultural evolution has historically lagged by decades behind conventional evolutionary theory. There was a revolution in the 1970s in which group selection fell out of fashion, and kin selection became much more popular. A parsimonious explanation of the penchant for group selection in the social sciences is that the field of cultural evolution has yet to go through this transition. The significance of relatedness between memes and memeplexes has yet to be fully appreciated.
They deny that evolution can be usefully viewed as an optimization process, saying:
Thus, evolution does not, in general, lead to the maximization of inclusive fitness or any other quantity.
That's complete nonsense: Allen, Nowak, and Wilson don't know what they are talking about. Optimization models are perfectly general - and can model any dynamical system.
The guts of their paper is a criticism of the application of Price's equation to inclusive fitness.
Their main beef is linearity. They are concerned that the 'adding' and 'subtracting' that goes on in the definition of inclusive fitness limits its generality. They write:
Inclusive fitness assumes that personal fitness is the sum of additive components caused by individual actions. This assumption does not hold for the majority of evolutionary processes or scenarios.
However, this linearity doesn't really cause problems - since you can approximate non-linear curves using a series of line segments.
Inclusive fitness is mostly concerned with the issue of whether some helping behaviour will evolve. It calculates whether the net selective effect of a behaviour on the frequency of a gene responsible for it (some of which may be copies in relatives of the actor) is positive or negative (relative to a set of alternatives) in the current environment. The math of inclusive fitness tells you whether the gene is selectively favoured. That's not quite the same thing as whether the behaviour will evolve (maybe it will drift into extinction) - but it's a good start. Nobody ever thought that the behaviours and genes involved had to not interact. Nobody thought that the rate of change of the frequency of the gene would be fixed over time. Maybe the environment will change in crazy non-linear ways in the future - and you'll have to redo your sums.
Basically, it isn't true that inclusive fitness theories assume that personal fitness is a linear sum of action fitness deltas. An action can change the environment (or the actor) in ways that affect the costs and benefits of future actions - and this is perfectly compatible with Hamilton's rule. It means that you may have to apply the rule more than once, is all.
Allen, Nowak, and Wilson's paper is like saying: I've got this crazy non-linear function, there's no way your linear approximation can match it. Except that: yes, there is - a series of linear functions can approximate any other function arbitrarily closely.
If you think that making this sort of objection is childish, I think that you're right - it is childish.
The paper says it was: "supported by a grant from the John Templeton Foundation". It seems like even more fuel for those who think that the Templeton Foundation is systematically distorting science.
Allen B, Nowak MA, & Wilson EO (2013) Limitations of inclusive fitness. Proceedings of the National Academy of Sciences of the United States of America
Kin selection theory can only be applied when organisms are influenced by their kin. However this happens at least once in every organisms' lifetime - when they are born.
Kin selection applies to calculations of parental investment and cases where there is parent-offspring conflict. Since all multicellular creatures have a parent-offspring asymmetry and provide some kind of resource bolus to their offspring, that means that kin selection applies to all multicellular organisms.
In other organisms, kin selection is also often relevant. Many microscopic organisms exhibit limited dispersal - and so are surrounded by their kin. If your kin are nearby, you should typically curb your pollution, share your food, and commit suicide if you are compromised by pathogens. Many parasites spend much of their time inside their hosts - where they are often surrounded by their kin.
Kin selection also applies to cultural variation - in the form of cultural kin selection - where it is similarly ubiquitous.
I think it is hard to argue that group selection has been anywhere near as productive as kin selection. However, group selection enthusiasts have been massively outnumbered over most of the recent history of the two ideas. Group selection defenders can claim that group selection has been less productive due to fewer man-hours expended on it.
The primary strengths of kin selection as I see them:
Kin selection makes extensive use of relatedness (a concept group selection enthusiasts rarely mention). This facilitates making quantitative predictions.
Kin selection has "Kin" in its title, which emphasizes the significance of kinship to the effect. Calling the effect "group selection" fails to to emphasize kinship. In practice this leads to applications where kinship isn't involved - and such applications are more often than not errors and mistakes.
The field is more mature, established and respectable.
I think that group selection appeals more to those who are attracted to the scientific fringes - in the hope of finding useful things there. That's all very well, but the the scientific fringes have a lot of junk science in them as well. Some of it is to do with group selection. Take care with it.
Kin selection applies to cultural variation. This is called cultural kin selection. It is an important topic.
Here's a brief history of the concept:
The observation that social and genetic kinskip were different in humans dates back to Lewis Henry Morgan in 1871.
Boyd and Richerson published one of the first articles on the topic in 1980 - connecting cultural kin selection and cultural group selection.0
An early modern article on the topic was written by Kristen Hawkes in 1983 and titled: "Kin Selection and Culture".
David Schneider's 1984 book "Critique of the Study of Kinship" argued that kinship was a different concept in different cultures, that people treating non-relatives as though they were kin was common - and that this reduced the significance of kin theories from biology when considering humans.
The idea of cultural kin selection appears in David Hull's (1988) book. David wrote:
Increasing one's conceptual inclusive fitness in science means increasing the number of replicates of one's contribution in the work of successive generations of other scientists.
...and...
science is a function of conceptual inclusive fitness.
Next, Francis Heylighen's (1992) paper: "Selfish Memes and the Evolution of Cooperation" stated:
The mechanism of kin selection can be extended to memes by redefining inclusive fitness as the fitness of a meme taking into account all its carriers. All individuals carrying the same meme can be viewed as relatives or kin insofar as this meme is concerned. Hence, the tendency to be altruistic towards offspring or close kin that follows from genetic evolution can be generalized to altruistic tendencies towards members of the same memetic or cultural group. The explanation for ultrasociality in the social insects on the basis of genetic inclusive fitness can be readily transposed to the emergence of ultrasociality in humans on the basis of memetic inclusive fitness.
He went on to explain how memes exploited kin-altruism towards relatives to redirect benefits towards cultural relatives:
many ethical systems explicitly refer to the ideal of "fraternity", and sometimes members of the same cultural group (e.g. monks or Freemasons) are supposed to call each other "brother". Though these are not brothers in the biological sense, the meme attempts to harness the innate tendency to behave altruistically towards kin and to use it for purposes different from the increase of genetic inclusive fitness.
Around the same time, Paul Allison published a series of papers on the topic - most notably "The Cultural Evolution of Beneficent Norms".
Heylighen revisited the idea of cultural kin in 1997, writing:
The genetic argument for altruism towards individuals carrying the same genes (kin selection) generalizes to altruism towards carriers of the same memes.
In 1997, David Hales wrote in "Memetic Evolution & Suboptimisation":
I now want to propose that two previously described extensions to basic natural selection can be applied subtlety within a meme framework: kin altruism and group selection.
There's a paper by John Evers (1998) called "A justification of societal altruism according to the memetic application of Hamilton's Rule", which attempts to directly adapt Hamilton's Rule to memetics - based on the idea of the "fraction of shared memes". John doesn't really go into the difficulties associated with this idea, though.
Also in 1998, "Cultural Software" by J. M. Balkin came out. Balkin clearly understood cultural kin selection, writing:
We often see people energetically promulgating their memes in the forms of beliefs, behaviors, artifacts, and customs while struggling with others who resist or disagree. Just as competition between biological kin groups can lead to strife, so can competition between cultural kin groups.
...and...
Just as individuals have varying degrees of genetic kinship, they also have varying degrees of memetic kinship. The two forms of kinship are cross-cutting: people can have many of the same memes even if they are completely unrelated. If the analogy to evolutionary arguments about kin-based altruism holds, then we would predict considerable altruistic behavior between people with lots of similar cultural software-for example, people of the same religion or culture, teachers and students, members of the same fraternity or club.
In 2000, David Hull showed that he understood the idea that kin selection extended into the cultural realm, writing: "One final example of similar processes operating in biological and memetic change is kin selection". He went on to give an example:
In science, scientists also distinguish between kin and non-kin, but the relevant genealogy is conceptual. The issue is not who holds similar ideas but who is conceptually connected to whom. The best way to increase the likelihood that you will be a successful scientist is to work under a successful scientist (Hull 1988).
Momme Von Sydow in a 2000 thesis wrote:
Corresponding to kin selection on the biological level, memes might ‘egoistically’ support their relatives in the same brain or in different brains, if they were identical (or similar enough). One might formulate a mathematical inequality analogous to Hamilton’s, where memes would replace genes.
In 2001, an important paper on tag-based cooperation came out - by Riolo, Cohen and Axelrod. It was followed by a large number of other papers on the topic. Riolo had come up with the idea much earlier - and had performed computer simulations of tag-based cooperation in 1992.
In 2001, Karl Sigmund and Martin Nowak, pointed out that cultural tag-based cooperation was a form of cultural kin selection.
Gordon Rakita (2003) rechristened cultural kin selection "kith selection". The term "kith selection" is kind-of cute - but it seems as though it is unnecessary jargon - and it doesn't fit in very well with "cultural group selection".
The idea of "cultural kin selection" appears again in Francis Heylighen's (2008) article: "Cultural Evolution and Memetics" - where he says:
Horizontal transmission of cooperation norms solves this problem, since the members of a cultural group are all memetically related to each other, sharing their memes rather than their genes. Therefore, cultural kin selection will extend to all members of the group (Evers, 1998). This entails a selective pressure for memes to support the fitness of the whole group of their carriers, e.g. by promoting cooperation.
In 2009, Gordon Rakita spelled out some of the implications of the idea of "kith selection" in detail. He started out by introducing a class of behaviours that kin selection acting on genes appears to explain poorly:
Kin selection does not seem to offer a suitable explanation for instances in which soldiers give their lives for their comrades. The conditions necessary for the model are not met, for example, soldiers are rarely genetically related. Given this, how are such acts to be explained?
He then offered a fine explanation - involving selection acting on memes:
A meme that encourages a soldier to throw himself on a grenade to save his comrades will negatively impact its future prospects of being transmitted to others. (This behavior is similar to that of the worker ant who forgoes reproduction in favour of the reproductive potential of the queen.) However, if the soldier’s comrades also carry the meme for grenade smothering, then the soldier’s behavior will indirectly impact the future replicative success of those other copies of the altruistic meme. This mimetic model of kin selection I term kith selection, kith being unrelated but like-minded acquaintances
Recently Mark Pagel's Wired For Culture book offered an explanation of human cooperation in terms of cultural "green beard" effects. In the process, he pointed out that such green beard effects were a kind of kin selection.
Group selection advocates often cite laboratory studies on breeding chickens to maximise egg production.
What they don't mention is that the birds involved were housed by sire family, were close relatives - and the whole process was conceived at the time in terms of kin selection. Check out this quote:
Craig (1982) advocated kin selection, in which pullets are housed in cages together as families with mean performance used as the criterion of selection. He hypothesized that families that perform best tend to have those physiological and behavioral characteristics most appropriate for group well-being and productivity. Muir (1985), Craig and Muir (1993), and Muir (1994) hypothesized that kin selection would favor cooperative tolerant behavior and concluded that selection on family means, when families are kept together as family groups, provides a method of improving traits in which behavioral interactions influence overall well-being and productivity.
The challenge for group selection has always been distinguishing itself from its widely-accepted rival, kin selection. The case of the chickens surely represents a failure to do this.
I re-read the Dawkins essay "Gods Utility Function" recently. Dawkins offers an argument against high-level selection in it. However, the article gives a string of dodgy examples. He argues that tree trunks are wasteful:
Why are forest trees so tall? Simply to overtop rival trees. A “sensible” utility function would see to it that they were all short. They would get exactly the same amount of sunlight, with far less expenditure on thick trucks and massive supporting buttresses. But if they were all short, natural selection couldn’t help favoring a variant individual that grew a little taller. The ante having been upped, others would have to follow suit. Nothing can stop the whole game escalating until all trees are ludicrously and wastefully tall. It is ludicrous and wasteful only from the point of view of a rational economic planner thinking in terms of maximizing efficiency. But it all makes sense once you understand the true utility function-genes are maximizing their own survival.
This is wrong. Very tall tree trunks are found in the most efficient dissipating regions of the earth: rain forests. These extract far more energy from incident sunlight that the grasslands that resemble the type of ecosystem which Dawkins is describing as moreefficient. Those tree trunks pay off in efficiency terms. If you have a flat surface, it heats up - and the incident energy radiates back into space. You need a lot of surface area to allow water evaporation to keep you cool. The depth produced by tall trees helps to provides such a surface.
In the essay, Dawkins also describes elephant seal sex ratios as "inefficient":
The sex ratio-the proportion of males to females-in wild populations is usually 50:50. This seems to make no economic sense in those many species in which a minority of males has an unfair monopoly of the females: the harem system. In one well-studied population of elephant seals, 4 percent of the males accounted for 88 percent of all the copulations. Never mind that God’s Utility Function in this case seems so unfair for the bachelor majority. What is worse, a cost-cutting, efficiency-minded deity would be bound to spot that the deprived 96 percent are consuming half the population’s food resources (actually more than half, because adult male elephant seals are much bigger than females). The surplus bachelors do nothing except wait for an opportunity to displace one of the lucky 4 percent of harem masters. How can the existence of these unconscionable bachelor herds possibly be justified? Any utility function that paid even a little attention to the economic efficiency of the community would dispense with the
bachelors. Instead, there would be just enough males born to fertilize the females.
Unfortunately for this argument, the surplus of males helps the population weed out parasite-ridden and mutation-loaded individuals. A population with fewer males would mean males of lower quality breeding - with the quality of individuals cumulatively declining over the generations. Dawkins' economic argument about how the makes are redundant is simply mistaken. The deaths of the failed males helps the population to adapt by marking out the adjacent gene-space that represent reduced quality - helping the population flee from its parasites and decrease its mutational load.
Perhaps it is possible for trees to be too tall - and for there to be too many elephant seal males. However tall trees and numerous male elephant seals proves very little.
Of course, the basic theme of this essay - that "the true utility function of life, that which is being maximized in the natural world, is DNA survival" is hopelessly mistaken.
Dawkins' argument against multi-level selection in this essay is also incoherent.
One thing I learned from reading "Social Bonding and Nurture Kinship" was that there was a debate over whether kin recognition and other adaptations related to kin selection are really all that different. I had previously absorbed (from the literature on the topic) the idea that "kin recognition" was a subset of "kin discrimination". Looking at another individual and seeing that they are similar to you and other family members is called "kin recognition". Helping nestmates (regardless of whether they resemble you) is called "kin discrimination".
However, another camp holds that kin recognition is what kin selection does. From this perspective dividing "kin recognition" from "kin discrimination" isn't carving nature at the joints. It's trying to make a distinction where there isn't really much of a difference:
I suggest that group or individual recognition systems might well qualify as kin recognition regardless of the cues used, provided that the groups or individuals so recognised are routinely kin and that the fitness benefits associated with recognition typically flow among kin. Grafen's insistence that kin recognition be a function of genetically based cues and that no individual or group recognition system should qualify excludes many recognition systems that may well function to associate kin in a fitness enhancing context. Kin recognition is an inherently functional concept and any definition or restriction that relies too much on a particular mechanisms or source of cues is problematic. (Stuart 1991).
I've warmed up to this. "Kin recognition" and "kin discrimination" is too much terminology for what is basically one type of effect. "Kin recognition" is the more popular term (by a factor of five). The "kin discrimination" category is surely the more useful one. I think we should just refer to "kin discrimination" as "kin recognition".
Kin selection theory raises the possibility that organisms may use variation between them as clues to relatedness - and direct cooperative behaviour preferentially towards perceived relatives. This effect is sometimes referred to as "kin recognition".
However, this method of identifying relatives depends on the existence of population-scale variation. Population bottlenecks can destroy such variation - and may promote cooperation.
Similarly, the founder effect might also produce local regions with little variation.
These effects have been demonstrated experimentally:
In theory, kin selection should mostly produce adaptations that work on relatedness cues that dynamically take overall population similarity into account. However, kin selection effects must work by manipulating development. Cruder measures of detecting similarity and relatedness will often be employed in practice.
Humans are among those species that have experienced a relatively recent population bottleneck - in the form of the Toba catastrophe. It is intriguing to consider the scale of the resulting increased levels of cooperation between humans that might be the result of this.
What do you get when you put two new theories together?
In the case of cultural evolution and group selection, what you seem to get is a big scientific muddle. Let me explain:
Many modern cultural theorists treat culture as a second inheritance channel that affects and modifies human phenotypes (extended phenotypes in the case of artifacts). This is instead of modeling cultural symbionts as separate lineages with distinct phenotypes of their own. For more on this see Against the extended genotype
This perspective has led to multiple claims that group selection is responsible for various features of human
culture:
There are two separate problems with this sort of literature:
The first is that typically no evidence is presented to show that these memes are deleterious within groups. If you count the spread of memes that are simply advantageous as a form of "group selection", then all widespread cultural phenomena qualify and the term becomes meaningless. Group selection theorists should try harder to distinguish between group selection and byproduct mutualism - which can both produce "groupish behaviour", but by independent mechanisms. It is true that some individuals benefit from polygamous marriage and anti-social selfishness, but other individuals are harmed, and if the balance is positive, group selection must compete with an obvious explanation: that some individuals are manipulating other ones using memes which have high average fitness and spread by perfectly ordinary natural selection.
The second problem is that - just because you can model a phenomena using multi-level selection and the Price equation, it doesn't mean that it makes sense to do so. Rather than observing that selection on human hosts and selection on memes operates on different scales - and reaching for the multi-level selection toolkit - what should happen is that the fact that humans and their memes are not geneaologically related should be noted, and they should be modeled as independent species using natural selection on humans and natural selection on memes. Multi-level selection is an unnecessary and confusing complication in such cases.
To illustrate, compare with the case of the smallpox virus. Smallpox germs wiped out many native Americans. The smallpox scabs are an aspect of individual human phenotypes. They are deleterious to individual humans (they kill people). However has historically been advantageous to human *groups* to have the smallpox phenotype - since this trait obliterated many competing tribes of humans (e.g. see American history). I think that few would advocate multi-level selection modeling in this case. There are two distinct types of entity involved here: humans and the smallpox virus. Natural selection operates on them more-or-less independently. Group selection brings no enlightenment and much confusion to this situation. The idea that smallpox scabs are traits which spread because they are deleterious to individuals and advantageous to groups is just a misleading and bad way of looking at the situation. It could equally well have been true that smallpox scabs were deleterious to human individuals and human groups - in which case it could still spread for its own "selfish" reasons. Group selection would then have been a red herring in explaining the spread of smallpox.
What holds for smallpox virus scabs holds for many kinds of memes. "Gun" memes result in people dying just as surely as in the case of the smallpox virus. Like smallpox, guns killed many native Americans - and gun memes provide broadly-similar group level advantages. You can model the spread of gun memes using multi-level selection and the Price equation. It is an unenlightening and confusing thing to do, but you can do it.
Its much the same with monogamy memes and cooperative memes. Memes don't interbreed with humans - they are more like separate species. So, you can forget about multi-level selection and just apply ordinary natural selection models to the humans and the memes - and have a much cleaner, neater model. Since human genes and memes are more like different species, it is an unnecessary source of complication and confusion to muddle them together in a unified multi-level selection model - just because they influence the same phenotypic traits sometimes.
It's worth sorting this muddle out - partly because memes exhibit their own kin/group selection dynamics, which can be important and significant. This is where genuine kin sleection/group selection operate in the cultural realm. If "cultural group selection" gets turned into a meaningless catch-all term for cultural phenomena which spread despite being deleterious to human hosts, it risks losing the possibility of performing useful work in other contexts.
References
Henrich J, Boyd R, & Richerson PJ (2012). The puzzle of monogamous marriage. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 367 (1589), 657-69 PMID: 22271782
The first category in this list represents kinship - or relatedness. It could be one organism
copied from the other, or that both copied from a shared source. This post
will argue that convergent evolution can be usefully seen as a type of kinship
or relatedness - that the first two categories are fundamentally similar -
and that the theory of kin selection applies to both of them.
In some cases, it is obvious that similarity between non-kin is due to copying. For example,
many unrelated insects resemble wasps. Bees are the best-known example - but many other
creatures have adopted the same colouring scheme. This is widely known as "mimicry".
This "convergence" of appearance seems fairly clearly due to information being
copied via vision systems of predators and enemies into the genes of insects.
Similarity involving camouflage colouration also plainly involves copying.
Many female birds are a mottled dirty brown colour. They are copying
colours from environmental backgrounds - in order to blend in.
They often look similar because they are copying from similar
environments.
So: some convergent evolution plainly involves copying - in the sense described in detail
here. However, the thesis here is that all convergent evolution involves copying. Let's look at a few more-challenging cases:
Birds and bats both have wings. They resemble each other - without being
closely related. Their last common ancestor had no wings and could not fly. What's
being copied here? Here, the copied information involves some aspects of a shared
environment, and some things derived from physical laws.
Marsupials and mammals share many common features - and it is widely agreed that many
are the result of convergent evolution. However, they evolved on different continents -
making copying difficult. What was copied here? Again, shared environments resulted
in similar selection pressures - and ultimately similar morphologies. The environments
were similar because they were on the same planet - with much the same fauna and flora -
due to shared descent.
Even convergent evolution based on the uniformity of nature can be regarded as being likely to
involve copying. Why are the laws of physics uniform? The answer involves identity copying.
Either the laws of physics were copied from a shared source when now-distant parts of
the universe were once closer together - or physical uniformity is due to something
like a state machine that iteratively deals with all locations in the universe at each moment
in time. The latter case also qualifies as copying - since by definition, copying involves
information in one place being later found in multiple places.
The applicability of kin selection theory to cases of convergent evolution is
in line with Price's
approach to the issue - which is based on correlations between traits. Convergent
evolution produces such trait correlations - it is appropriate that kin selection also
deals with these.
"Kin selection" and "group selection" are common terms for the way that
organisms help organisms which are similar to themselves. I've
proposed the term "similarity selection". Another contender is
"clade selection". This term came from George C. Williams (1992).
Williams efficiently disposed of “group selection,”
which never recovered (except as a muddled version of kin selection).
But in Natural Selection: Domains, Levels and Changes (1992),
where he gathered many threads of thought, he developed the
important and superficially similar idea (foreshadowed in
Adaptation and Natural Selection) of “clade selection” to explain,
not “altruism” but macroevolutionary patterns of diversity and - as
I would put it - “the evolution of evolvability.”
"Clade selection" seems like better terminology than "group
selection" - in some respects. Like the term "kin selection"
it puts an explicit emphasis on relatedness. Group selection
only works - in the sense of producing group-level adaptations -
when relatedness is involved. So: why not use the term clade selection?
Alas, I think there's a good reason not to use the term "clade selection".
A clade consists of an organism and all its descendants. The problem
is with the "all". You can still have group selection that works
without it being clade selection - by the fitness delta involved
not affecting all the descendants.
The "all" in "clade" makes "clade selection" a confusing term. Not worthless,
perhaps, but I don't think I can endorse the term.
This is a pretty technical presentation by group selection advocate Rob Boyd on kin selection and the dust-up relating to it in Nature. It's slides-and-audio only (sorry).
Boyd calls Stuart West and Andy Gardner the "Inclusive fitness mafia" 31 minutes in.
He presents an objection to kin selection theory, calling Hamilton's rule "true but misleading" 50 minutes in. It seems like sour grapes to me.
Hi. I'm Tim Tyler, and this is a review of this book:
The Social Conquest of Earth by Edward O. Wilson
The book is about the rise of social organisms on the planet and how they have come to dominate their ecosystems. Wilson uses ants and humans as his main examples of the success of sociality.
It's aiming to be a popular book. There are, in fact, a few scientific references in the book, but if you're not careful, you won't find them until you get to the very end - where they are clustered together in a list.
The first 100 pages of the book is a history of the rise of humans. Alas, I'm one of those who often finds histories boring, so I had to push a bit through this section of the book. Then there's thirty pages on social insects. Then there's sixty pages on the theories of social evolution. This is really where Wilson comes unstuck.
Wilson launches into a misguided attack on kin selection theory. Obviously, Wilson has some understanding of kin selection. However, he doesn't have a sympathetic understanding of it, and instead seeks to destroy and discredit it. Alas, in order to do this, he has to set up a series of straw men and then vigorously attack them. This might be entertaining for some - but Wilson is a popular author, and here we run the risk of an eminent scientist confusing and misleading the next generation of students. About the best that can be said for Wilson's attack on kin selection is that it is embarrassing and half-baked. I noticed that, as if to illustrate that he doesn't really know what he is talking about, Wilson gives an incorrect definition of inclusive fitness, saying:
The inclusive fitness of the individual is its personal fitness, in other words the number of its personal offspring who grow up and have children of their own added to the effect its actions will have on the fitness of its collateral relatives, such as siblings, aunts, uncles, and cousins.
That definition is wrong. Those familiar with the topic will recognise that it's only two thirds of the correct definition. Wilson goes on to discuss the claimed equivalence of kin and group selection. He says:
Further there are mathematical difficulties with the definition of r, the degree of relatedness. These difficulties render incorrect the oft-repeated claim that group selection is the same as kin selection expressed through inclusive fitness.
After supposedly demolishing the decades of work by kin selection theorists, Wilson goes on to present his preferred alternative theory of social evolution - which is based on group selection and the idea of group-forming forces - where Wilson gives a "defensible nest" as his main example of the latter.
Unfortunately, Wilson's book is probably exhibit number one when it comes to showing how group selection leads to confusion about evolutionary theory. Wilson is evidently in a hopeless muddle about kin selection and group selection. It seems hard to avoid the conclusion that group selection lies at the root of the problem. While Wilson might have been trying to help the cause of group selection in his book, it seems more likely that it will be an embarrassment for group selection's proponents.
Group selection's main problem that it faces a well-armed opponent. Kin selection explains all the same things that group selection explains - and has been scientific orthodoxy for decades. Most modern group selection enthusiasts seem to have finally agreed that group selection makes no predictions not also made by kin selection. The main case for group selection these days seems to amount to the idea that it represents a stimulating alternative perspective. High-level selection can be a form of holism to counter kin selection's reductionism. Group selection models can be tractable in some cases where lower-level kin selection models are not. This is all reasonable: group selection does have a positive side. The problem with group selection is that historically its use has often led to junk science. People tend to invoke benefits to group willy-nilly - without doing the math, or even sanity-checking their explanations. To many scientists, group selection explanations thus often look like a poor substitute for actually understanding what is going on. Many explanations based on group selection, on closer examination, have turned out to be wrong or misleading. In the cases group selection is useful, kin selection is often more useful. SO, group selection has its merits - but needs a health warning.
Wilson claims that humans are a eusocial species - even though we mostly lack one of the primary traits associated with eusociality - namely reproductive division of labour. Wilson doesn't explain that his use of the term is unorthodox and controversial.
Wilson's examples of eusociality are all from animals. He lists beetles, aphids, thrips, shrimps, mole rats and humans. However there are other examples of social behaviour among microbes, plants and fungi. In particular multicellularity is a common form of eusociality. The examples of its evolution illuminate the topic of social evolution. However, Wilson makes no use of these examples.
Wilson attempts to apply the study of animal social evolution to humans - and this leads him to the kin selection (or group selection) axis. While important, this leads to a pretty myopic perspective on human social behaviour, which is heavily influenced by reciprocity, reputations, byproduct mutualism - and manipulation of humans by memes. You can't really coherently discuss human social evolution without casting your net wider than this.
Lastly, the book ends up with a review of cultural evolution in humans. In the 1980s Wilson was a pioneer in applying biology to human culturally-transmitted behaviour, writing two books on this topic. Wilson at least understands that culture evolves - and that memes coevolve with genes. However, his older books promoted a pre-memetics perspective, in which everything boiled down to DNA genes. Checking to see if Wilson had moved on 40 years later, I was surprised to find that little had changed. He cited his old work, and trotted out much the same theory that he advocated back in the 1980s. This idea has been totally superseded in the mean time by the application of Darwinian evolutionary theory directly to cultural variation - along the lines pioneered by Richard Dawkins in 1976. Wilson's discussion of this work is confined to one paragraph. He chastises the researchers involved for paying insufficient attention to the interactions between genes and memes - and then launches into a laundry list of such interactions.
This hopeless treatment of cultural evolution might not matter - but this is one of the few books addressing the topic that I regularly see in bookshops. This might be the first exposure to the topic for many. A teaching opportunity has been squandered, because Wilson is stuck back in the 1970s on the topic and has failed to grok most of the more recent literature.
In the book, Wilson expands on his old claim that genes keep memes on a leash by presenting an argument against the possibility of a memetic takeover. He says:
I am further inclined to discount the widespread belief that robotic intelligence will overtake and potentially replace human intelligence.
What are Wilson's reasons? He goes on to give them, saying:
To advance from robot to human would be a task of immense technological difficulty. But why should we even wish to try? Even after our machines far exceed our outer mental capacities, they will not have anything resembling human minds. In any case, we do not need such robots, and we will not want them. The biological human mind is our province.
Wilson's objection seems simple-minded to me. He gives me no reason to think that he knows much about machine intelligence or futurism. He's just a famous biologist speculating on something about which he knows little. The idea that people will lack motivation to build superhuman intelligent machines just seems daft to me. The motivation is obvious.
So: this is not a good book. It is quite readable, but it's full of outdated or wrong science. Readers should be warned that, while Wilson speaks authoritatively, he doesn't really know what he's talking about in many places.
Hi. I'm Tim Tyler, and this is a review of this book:
Darwinism and Human Affairs by Richard Alexander
The book was published in 1979. The contents represent what we would now call sociobiology. Unfortunately, this is the bad kind of sociobiology that tries to trace everything back to DNA genes.
The book offers a perspective similar to that of E.O. Wilson - who published similar works at around the same time.
I read it mainly in order to understand Richard's perspective on human culture from that era.
The book predates our modern understanding of cultural evolution and instead presents a view of culture based on DNA genes.
In a section titled: A comparison of organic and cultural evolution, Richard breaks cultural evolution down into inheritance, mutation, selection, drift and isolation. He then reviews existing literature relating to the modern perspective on cultural evolution citing the views of Dawkins, Durham, Cloak, Cavalii-Sforza, Feldman, Richerson and Boyd on the topic. This might all sound good. However, the section then goes on to dismiss this material, saying:
regularity of learning situations or environmental consistency is the link between genetic instructions and cultural instructions which makes the latter not a replicator at all, but in historical terms a vehicle of the genetic replicators.
Having thus dismissed the Darwinian view of culture which does so much useful work in modern times, Richard offers in its place the idea that cultural information is a persistent part of the environment that is influenced and manipulated by genes. This is true - as far as it goes. However, these days, we know that you can get a lot of mileage out of the idea that cultural variation evolves in a very similar way to the way genetic variation evolves - complete with selection, adapatation, drift, recombination, kin selection, gradualism and heritage constraint. Richard's model is not exactly wrong, it just isn't as helpful as it could be. Because culture evolves, evolutionary theory has a rich array of tools and models which can be directly applied to cultural variation without much modification. If you fail to recognise that, you don't get to use these tools and models.
It is rather frustrating for the reader to see the author toy with the correct theory - and then abandon it. On a more positive note, it was probably good to have the position that everything boils down to genes clearly articulated - to give critics something to argue against.
Most of the rest of the book is concerned with explaining various types of human behavior in terms of DNA genes. Richard looks at nepotism, incest avoidance, xenophobia and various other aspects of human affair with a biological basis. He engages with anthropologists critical of biological approaches. While he makes most of them look stupid, some of their objections make some sense retrospectively. One thing they were critical of was the over-application of kin selection theory. Looking at the kin selection being advocated back then, only kinship between DNA genes was used - and all other kin-like relationships were classified as fictive kinships, with the organisms involved being manipulated and fooled into thinking their colleagues are their kin. This now looks like a terribly impoverished kind of kin selection, in the light of cultural kin selection. There's memetic kinship, as well as genetic kinship. The anthropologists didn't have the concept of cultural kin selection back then either, but at least they recognized that there was much more to kinship than relatedness between DNA genes.
Richard bashes group selection here and there in the book, but his story of how humans evolved revolves heavily around conflict between groups. Because of this, modern group selection enthusiasts may find him an ally as much as an adversary. Here's a sound bite from the book on the non-equivalence of kin and group selection:
Despite the recent prominence accorded to the view that the maximization of inclusive fitness by helping the aggregate of one's relatives - or what Maynard Smith called "kin selection" is a kind of group selection, this is a misleading, if not erroneous view. Group selection thwarts the reproductive interests of individuals when these interests differ from those of the group. Kin selection is a way in which individuals further their genetic interests via other individuals who carry some of their genes.
While this might have sounded plausible at the time, it now looks wrong. Group selection has turned out to be a way in which heritable information perpetuates itself via promoting copies of itself in other group members.
The book finishes up with a look at justice and ethics. Richard says:
Arguments given above, and the cited references make it clear why I believe that evolution has more to say about why people do what they do than any other theory. In contrast, my answer to the question "What does evolution have to say about normative ethics, or defining what people ought to be doing?" is "Nothing whatsoever."
I think this is a cop-out. Evolutionary theory surely has more to say about this subject than any other branch of science. If intelligent people aren't supposed to use science to guide their ethics, then it isn't clear where they are supposed to be getting their ethical principles from. Richard says that he thinks that pain and pleasure are the basis of most normative ethics - but even if one accepts that, classifying these as "non-evolutionary" ignores the idea that the brain evolved, and the idea that the brain evolves over an individual's lifetime, using copying, variation, and selection in order to to seek its rewards. I don't think you can get away from evolutionary ethics this easily.
This is a nice book to read, but pretty out-of-date these days. However it is interesting to see part of the background that modern theories of culture and human evolution were developed against.
I was planning to publish this bibliography of cultural kin selection as part of my forthcoming "Memes" book - which will have chapters on cultural nepotism and cultural eusociality. However, several issues have resulted in me changing my mind: the URLs are more use on the internet - and a technical bibliography doesn't necessarily seem to be the best use of pages in a popular book.
Haun, D. B. M., & Over, H. (2013). Like me: A homophily-based account of human culture. [paywall] In P. J. Richerson, & M. H. Christiansen (Eds.), Cultural Evolution: Society, technology, language, and religion.
Looking at the science news, you might think that the news concerning kin selection has something to
do with group selection. However, that affair is a bit of a storm in a teacup, with
little real impact on the theory of kin selection.
I think the biggest impact on kin selection concerns the expansion of Darwinism's domain.
Kin selection was once only applied to genes. However, with the realisation that it also
applies to memes, individually-learned ideas, mimicry and other copied entities some
previously-unknown vistas have opened up for the theory.
The impact has been broadly similar to the discovery of alien life forms: new evolutionary dynamics let us know what is contingent on our local historical circumstances.
Is it true that kin selection's opponents are funded by the Templeton Foundation?
Jerry Coyne alleges this:
Finally, much of the work on group selection has been funded by the John Templeton Foundation, an enormously wealthy organization with an agenda to harmonize faith and science. The idea of group selection, with its spiritual and religious connotations—the process is often used to explain the prevalence of religion and societal harmony—is right up their alley. So the proponents of group selection monopolize not only the megaphones but the funding. In science, money talks.
Nowak's grant was ten million dollars - we are not talking about peanuts here.
The Templeton Foundation funding puts a strange slant on the issue. It is strange to think that some of
the skirmishes in the ancient battle between science and religion are now being shrouded in arcane
mathematics and published in top science journals.
 After the 2010 Nowak, Tarnita, & Wilson affair, Samir Okasha weighed in with a comment in Nature, effectively asking: why can't we all just get along?
After the 2010 Nowak, Tarnita, & Wilson affair, Samir Okasha weighed in with a comment in Nature, effectively asking: why can't we all just get along? You might think Nowak and Wilson would have had their fingers burned in the reaction to
You might think Nowak and Wilson would have had their fingers burned in the reaction to  Kin selection theory can only be applied when organisms are influenced by their kin. However this happens at least once in every organisms' lifetime - when they are born.
Kin selection theory can only be applied when organisms are influenced by their kin. However this happens at least once in every organisms' lifetime - when they are born. West Griffin and Gardner wrote a comparison of kin selection and group selection, titled:
West Griffin and Gardner wrote a comparison of kin selection and group selection, titled:
 Kin selection applies to cultural variation. This is called
Kin selection applies to cultural variation. This is called  Group selection advocates often cite laboratory studies on breeding chickens to maximise egg production.
Group selection advocates often cite laboratory studies on breeding chickens to maximise egg production. I re-read the Dawkins essay "
I re-read the Dawkins essay " One thing I learned from reading "
One thing I learned from reading "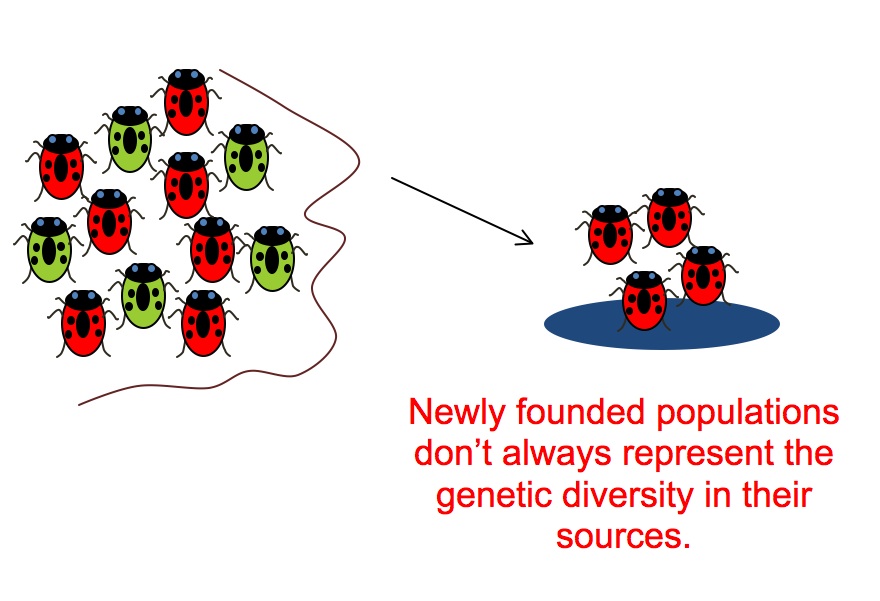 Kin selection theory raises the possibility that organisms may use variation between them as clues to relatedness - and direct cooperative behaviour preferentially towards perceived relatives. This effect is sometimes referred to as "kin recognition".
Kin selection theory raises the possibility that organisms may use variation between them as clues to relatedness - and direct cooperative behaviour preferentially towards perceived relatives. This effect is sometimes referred to as "kin recognition". What do you get when you put two new theories together?
What do you get when you put two new theories together? When organisms appear to be similar to one another there are several possible causes:
When organisms appear to be similar to one another there are several possible causes:

 "Kin selection" and "group selection" are common terms for the way that
organisms help organisms which are similar to themselves. I've
proposed the term "similarity selection". Another contender is
"clade selection". This term came from George C. Williams (1992).
"Kin selection" and "group selection" are common terms for the way that
organisms help organisms which are similar to themselves. I've
proposed the term "similarity selection". Another contender is
"clade selection". This term came from George C. Williams (1992). Looking at the science news, you might think that the news concerning kin selection has something to
do with group selection. However, that affair is a bit of a storm in a teacup, with
little real impact on the theory of kin selection.
Looking at the science news, you might think that the news concerning kin selection has something to
do with group selection. However, that affair is a bit of a storm in a teacup, with
little real impact on the theory of kin selection. Is it true that kin selection's opponents are funded by the Templeton Foundation?
Is it true that kin selection's opponents are funded by the Templeton Foundation?